
古生物學研究之中,最有樂趣的部份之一,就是發現並命名一個全新的物種。在過往生命歷史中,有許多我們不知道的生物存在,因此相對於現在還活蹦亂跳的生物而言,古生物並明相對容易而機會也大得多。而大部分的分類及命名等,都是從在我們眼前還活生生的生命開始。
每年眾多新命名的化石物種之中,今年(2016年)的這一個新名字或許會讓人特別覺得興趣盎然:忍者海豚!
木村(T. Kimura)和巴恩斯(L. G. Barnes)今年在日本群馬縣立自然史博物館研究報告(Bulletin of Gunma Museum of Natural History)的一篇文章中命名了三種新的海豚,而其中一個新屬新種的名字就被叫為:氏原忍者海豚(學名為Ninjadelphis ujiharai)。

挖掘古生物化石 命名新物種
如同其他被命名的化石物種一樣,氏原忍者海豚目前也就僅有這麼一件化石標本[1],是由氏原溫(Ujihara Atsushi)於1998年的時候,在日本三重縣(Mie)的伊賀市(Iga)發現的。
伊賀市是著名的忍者出沒地點之一,所以木村決定以化石的新名字紀念發現化石的氏原溫及其而對古生物學的貢獻,另外再以伊賀市著名的忍者來標記這件化石,也因此就有了氏原忍者海豚(Ninjadelphis ujiharai):ninja就是忍者,delphis為海豚,而ujiharai就是氏原溫了。
氏原忍者海豚和木村及巴恩斯文章中的另外兩種新化石物種:俄勒岡格德特海豚(Goedertius oregonensis)和唐娜長吻鯨(Zarhinocetus donnamatsonae),都是被歸類在同一個科:奇異海豚科(Allodelphinidae,allo有奇特或奇異的意思,而delphinidae為海豚科,所以翻為「奇異海豚科」)。從名稱來看,Goedertius是紀念格德特夫婦(J. L. Goedert和G. H. Goedert)長期在美國西岸尋找化石,並且將找到的化石捐獻給博物館,因此對於海洋哺乳動物化石的研究有極大的貢獻,而oregonensis是指出這化石發現的地點在美國的俄勒岡(Oregon),因此翻成俄勒岡格德特海豚。
另一名成員為唐娜長吻鯨(Zarhinocetus donnamatsonae),希臘文中的za有極度或非常的意思,rhino在希臘文為鼻子或吻部,cetus為拉丁文中的鯨魚,而donnamatsonae是紀念去年(2015年)過世的唐娜・馬森(Donna M. Matson)還在世時,會捐獻資金提供給巴恩斯工作的洛杉磯自然史博物館(Natural History Museum of Los Angeles County)進行海洋哺乳動物化石的研究,因此翻成唐娜長吻鯨。
忍者海豚和白鱀豚是好朋友
有趣的是,氏原忍者海豚所在的奇異海豚科,其實在更高一階的生物分類類群裡為:河豚!
我們在〈河裡的海豚還是海裡的河豚〉大概簡略地介紹了現生四種被稱為「河豚」的海豚(其中,白鱀豚或許不該再被歸類在現生物種裡了,因為基本上已經滅絕了。)及他們相關的化石和演化史。而奇異海豚科雖然已經完全絕滅,沒有現存的後代,但就骨骼形態來看,他們和現生恆河豚科(Platanistidae)的恆河豚有著相當親近的關係,因此目前被歸類在恆河豚超科(Platanistoidea)裡。
雖然現生的恆河豚超科目前只剩下生存在恆河及印度河流域的恆河豚,但從化石紀錄來看,恆河豚超科曾經一度在全球的海洋中大放異彩。除了現生恆河豚的生存危機之外,恆河豚超科不論是就生物分類所涵蓋的物種類群,或是地理分布來看,都是值得我們一窺他們繽紛的演化史。
即使有很多的化石物種還是讓我們感到很困惑,到底我們該將他們放在哪一個生物類群裡,還是備受爭議。需要更多的研究,最好是找到更多完整的化石,然後詳細的解析他們彼此間的形態結構。而恆河豚超科除了現生恆河豚的恆河豚科以及〈河裡的海豚還是海裡的河豚〉裡提到的懷依派特豚科(Waipatiidae)還有我們這裡談的,除了忍者海豚所處的奇異海豚科之外,至少還有另外四大類及一些還不清楚該放在這恆河豚超科目前7個科之中的那一個科的化石物種。這些目前「暫時」被歸類在恆河豚超科的類群,仍有爭議。而這四大類及其他分類層級未定的種類所隱含的故事瑣碎、繁雜,之後再另外找機會談。
光就我們談到恆河豚超科的兩個化石類群:懷依派特豚科和奇異海豚科的地理分布,大概就可以體會到他們在全球中廣泛的分布,懷依派特豚科的成員主要生活在南半球,而奇異海豚科則佔據了北太平洋的海域。
補完化石地圖 揭開生命歷史
而化石另一個有趣的地方就是,如果我們沒有去找,永遠都不會知道我們研究的化石類群曾經在哪一個地區出沒過。就像是皮亞努契(G. Bianucci)和他的研究團隊於2011年的時候,在地質生物期刊(Giobios)的一篇研究文章中,發表了從地中海馬爾他島(Malta)所發現的一些2000多萬年前的化石,很有可能是懷依派特豚科的成員,但因為這些化石並沒有很完整,所以確切分類的歸屬還有待更多新的化石來確認,但這化石紀錄也給了我們一些暗示,指出懷依派特豚科的分布,很有可能不止侷限於南半球。
再回頭來看奇異海豚科,同樣的,新的化石發現,持續的更新我們對於過往生命歷史的了解。在今年(2016年)之前,奇異海豚科只包含了三個物種,而木村和巴恩斯的文章一口氣就替奇異海豚科新增了三個成員,以數量來看,等於就是馬上倍增了!
不論是先前的三名成員或是木村和巴恩斯的三個新成員,全都是生活在太平洋比較低緯度的地區。但同樣在今年,博爾司馬(A. T. Boersma)和派恩森(N. D. Pyenson)也剛在同儕研究期刊(Peer J)命名了另一名奇異海豚科的成員:亞卡塔加北臉海豚(Arktocara yakataga)。arktos在希臘文中為北方的意思,cara在拉丁文中有臉的含義,因此Arktocara為:「北方的臉」,指出這件化石是在高緯度的地區所發現的,再加上目前唯一發現的化石標本[2],只有臉的部份,所以博爾司馬和派恩森用北方的臉來取名;而yakataga(亞卡特加)為特林吉特語(Tlingit)中,化石所發現的地點。所以翻成亞卡塔加北臉海豚。
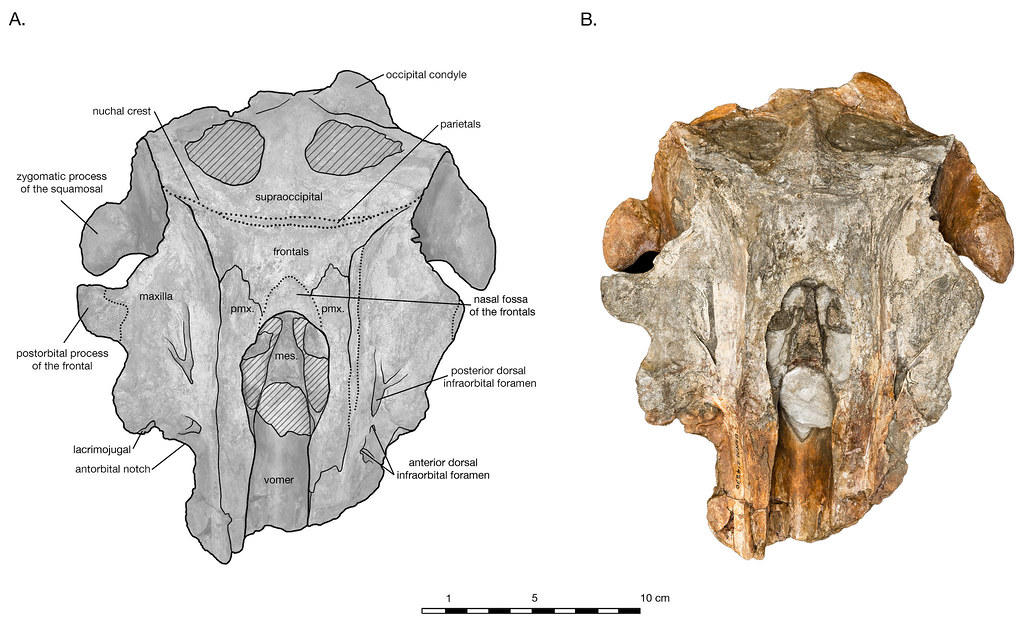
亞卡塔加北臉海豚是在阿拉斯加的東南邊發現的,而這化石不只是以地理分布來看是生活在最北邊的奇異海豚科內的成員,也是目前所有奇異海豚科成員中最古老的物種!
其他目前已知的6種奇異海豚科的成員,當然包含了大家可能很感興趣的忍者海豚,氏原忍者海豚的生存年代大約在1700 萬年前,都是在地質時期的中新世(Miocene。中新世的時間點約在2300萬年至500萬年前這區間)所發現的,但亞卡塔加北臉海豚是唯一從漸新世(Oligocene。介於3300萬年至2300萬年前)發現的奇異海豚。
雖然就亞卡塔加北臉海豚發現的地層來看,我們只能知道他們大概生存於2900萬年至2400萬年前這段時間,這中間橫跨了約500萬年的時候,目前還沒有更進一步的資料可以精確的縮小這時間範圍。
像這樣500萬年的不確定性,在古生物學中算是相當習以為常的事情,就現況而言,這樣的誤差並不是我們當下可以解決的,所以我們也算是可以和這種數百萬年的不確定性和平相處,但在有些討論的議題上,不只會讓我們有點困擾,也會引起不同古生物學家之間的議論。某種程度上來說,這樣的不確定性也是古生物學研究中很大的樂趣之一,因為這樣會驅使我們不斷地思考及去尋找更多的化石證據等來佐證各自的觀點。
尋找真相的化石拼圖
像是如果要討論一個類群的起源、或是他們遷移的過程及路線等等,有確切或是清楚的相對的時間就很重要了:我們常常都會以一個類群所發現最早的化石地點,當作是這一個研究類群的起源中心。
前面提到都是同樣屬於恆河豚超科的懷依派特豚科和奇異海豚科,一個主要佔據了南半球的海域,另一個主要生活在北半球的太平洋中。
就先前的狀況來看,遨遊在南半球的懷依派特豚科的成員都出現在時間點較早的漸新世時期,而所有發現的奇異海豚類群都生活在時間較晚的中新世(先不考慮恆河豚超科內的其他成員),雖然我們還沒有確定他們彼此間確切的演化關係,像是懷依派特豚科裡的成員是不是有包含了奇異海豚科的祖先,但從這樣的化石分布來看,我們大概可以大膽的假設:恆河豚超科是起源於南半球,並在漸新世/中新世交界的時間點左右,大約在2300萬年前,從南半球一路往北,佔據了北太平洋的海域,並且演化成我們稱呼他們為奇異海豚科的類群。
但是,僅僅單一個化石標本,一個從現今阿拉斯加的漸新世的地層中發現的亞卡塔加北臉海豚就可以指出這樣的假設是會有問題的:因為生活於北半球高緯度的亞卡塔加北臉海豚和南半球的懷依派特豚科同樣生活在漸新世的晚期,而兩個類群確切生活的時間點的不確定性都橫跨了數百萬年,所以其演化的先後順序,還需要更多的化石,及其他研究(例如準確的地層定年),才能幫助我們釐清他們多變的演化史。
等待發掘的台灣古生物
到這裡,或許大家多少可以感受到化石的威力及魅力。
以科學研究及想要瞭解過往生命歷史的角度來看,一個化石可以讓我們重新思考及測試先前的假說;而從本土或是人文的視角來思考,當一個新發現的化石,透過科學研究確定了隸屬於一個新物種,命名從當地發現的新物種的權利及機會,或許都會不斷的加強生活於該地區民眾的認同感及自信心。同時,收藏了新物種模式標本的博物館也持續的提高國際間的能見度:因為通常都只有那麼一件化石標本,全世界研究該類群的研究員如果想研究或是重新審視該化石,一定是需要親自到該博物館去親眼仔細的觀察該標本。
什麼時候,我們也會願意投入一定的心力、資源,發掘並詳細的研究在台灣還沒被發現的新化石物種,一方面可以利用從台灣發現的新化石來改寫一部份全球生命的演化紀錄;另一方面,這在過程所發現的新物種,也可以取上有當地特色及有趣的名字,像是我們今天認識的「忍者海豚」。

【備註】
- 氏原忍者海豚目前僅有的一件化石,收藏於群馬縣的自然史博物館。
- 收藏於美國史密遜自然史博物館National Museum of Natural History, Smithsonian Institution)
※ 本文與 行政院農業委員會 林務局  合作刊登
合作刊登




